KrytykaTeorii Poliwagalnej:kompleksowa analiza
dr Stephen W. Porges
Wybitny naukowiec uniwersytecki, Instytut Kinseya
i Uniwersytet Indiana
W skrócie
Teoria Poliwagalna wniosła biologicznie ugruntowane ramy pozwalające zrozumieć regulację układu autonomicznego, poczucie bezpieczeństwa oraz zaangażowanie społeczne, dostarczając wiedzy wykorzystywanej w badaniach i praktyce klinicznej, neuronauce, psychologii, medycynie i edukacji. W miarę jak teoria zyskiwała szerszą rozpoznawalność, stała się także przedmiotem krytyki zarówno w środowiskach akademickich, jak i w przestrzeni publicznej. Na tej stronie przedstawiono zwięzłe, oparte na dowodach wyjaśnienie kluczowych twierdzeń Teorii Poliwagalnej, jej zakresu oraz podstaw naukowych, aby ułatwić rzetelną ocenę poprzez odwołanie do źródeł pierwotnych i współczesnej neurofizjologii.
Opublikowane krytyki Teorii Poliwagalnej, w szczególności autorstwa Paula Grossmana oraz E. W. Taylora, wykazują powtarzający się wzorzec błędnych interpretacji i nieścisłości faktograficznych. Pomimo licznych wyjaśnień przedstawionych w literaturze recenzowanej (Porges, 2007b, 2022, 2023, 2025a, 2025b), krytyki te nadal wprowadzają błędy koncepcyjne zarówno do dyskursu akademickiego, jak i do praktyki klinicznej. Krytyka Grossmana z 2023 roku powtarza wątki wcześniej wysunięte przez Taylora, w dużej mierze opierając się na jego błędnych odczytaniach anatomicznych i ewolucyjnych, ujętych w retorycznie autorytatywnym stylu, który może wprowadzać w błąd czytelników nieznających literatury stanowiącej fundament teorii.
Oprócz publikacji recenzowanych, źródła skierowane do szerokiego grona odbiorców — w szczególności hasło w Wikipedii dotyczące Teorii Poliwagalnej — odegrały rolę w rozpowszechnianiu nieścisłości. Wpis ten wyraźnie przywołuje dwie krytyki: Neuhuber i Berthoud (2022) oraz Doody, Burghardt i Dinets (2023). Choć oba artykuły przeszły proces recenzji naukowej, ich publikacja w numerze specjalnym czasopisma Biological Psychology, redagowanym przez Grossmana, sugeruje redakcyjną zbieżność, która prawdopodobnie wpłynęła zarówno na ich ujęcie tematyczne, jak i koncepcyjne.
Krytyka autorstwa Neuhubera i Berthouda stanowi przykład błędnej interpretacji o charakterze anatomiczno-funkcjonalnym. Kwestionując funkcjonalne ujęcie dróg nerwu błędnego proponowane przez Teorię Poliwagalną, nie podważają oni podstawowych rozróżnień neuroanatomicznych — w szczególności oddzielenia grzbietowego jądra ruchowego nerwu błędnego (DMNX) od jądra dwuznacznego (NAmb), ani odmiennego stopnia mielinizacji ich włókien eferentnych. Pomijają jednak kluczowe założenie teorii: że System Zaangażowania Społecznego (SES) wywodzi się ze struktur łuków gardłowych w rozwoju embrionalnym oraz że komunikacja między kardiohamującym nerwem błędnym a SES ma w dużej mierze charakter pośredni, z wyjątkiem bezpośrednich połączeń obejmujących nerw trójdzielny. Wikipedia, streszczając tę krytykę, dodatkowo ją zniekształca — przedstawiając ją jako kategoryczne podważenie anatomicznego fundamentu Teorii Poliwagalnej, czego pierwotny artykuł nie twierdzi.
Podobnie krytyka autorstwa Doody’ego, Burghardta i Dinetsa błędnie przedstawia filogenetyczne ujęcie teorii. Autorzy kwestionują to, co interpretują jako sztywną dychotomię między „aspołecznymi gadami” a „społecznymi ssakami”. Tymczasem Teoria Poliwagalna jest wyraźnie zorientowana na ssaki i definiuje zachowania społeczne z perspektywy ssaczej. Podkreśla, że socjalność — w kontekście Teorii Poliwagalnej — ma swoje źródło w specyficznej dla ssaków innowacji autonomicznej, czyli brzusznym kompleksie błędnym, odpowiedzialnym za regulację Systemu Zaangażowania Społecznego. Choć gady mogą wykazywać adaptacyjne zachowania społeczne, są one mediowane przez odmienne systemy sygnalizacyjne i architektury neuronalne oraz pełnią inne funkcje ewolucyjne. W konsekwencji zachowania społeczne u gadów, choć ewolucyjnie adaptacyjne, nie są równoważne ze ssaczą socjalnością w rozumieniu Teorii Poliwagalnej. Zrównując analogię behawioralną z homologią neurofizjologiczną, krytyka pomija specyficzny zakres zastosowania teorii. Publiczne omówienia, w tym Wikipedia, dodatkowo wzmacniają to zniekształcenie, wyolbrzymiając znaczenie krytyki i pomijając nacisk Teorii Poliwagalnej na hierarchiczną ewolucję autonomiczną.
Co istotne, problem nie polega jedynie na tym, że te artykuły istnieją, lecz na tym, że zostały przefiltrowane przez platformę publiczną w sposób, który zarówno wyolbrzymia, jak i zniekształca ich treść. Ta wielowarstwowa błędna charakterystyka — ukształtowana przez osobiste uprzedzenia, a nie przez rzetelną analizę naukową — przyczynia się do rozpowszechniania wprowadzających w błąd informacji. Podkreśla to potrzebę oceny naukowej opartej na bezpośrednim kontakcie ze źródłami pierwotnymi, a nie na interpretacjach wynikających ze zbieżności redakcyjnej, selektywnego akcentowania czy wtórnego rozpowszechniania.
Aby wyjaśnić charakter tych błędnych interpretacji, w Tabeli 1 wskazano powtarzające się obszary błędów obecne w krytykach Teorii Poliwagalnej, przedstawiając sposób, w jaki się one przejawiają, oraz konsekwencje, jakie niosą dla trafnej interpretacji teoretycznej.
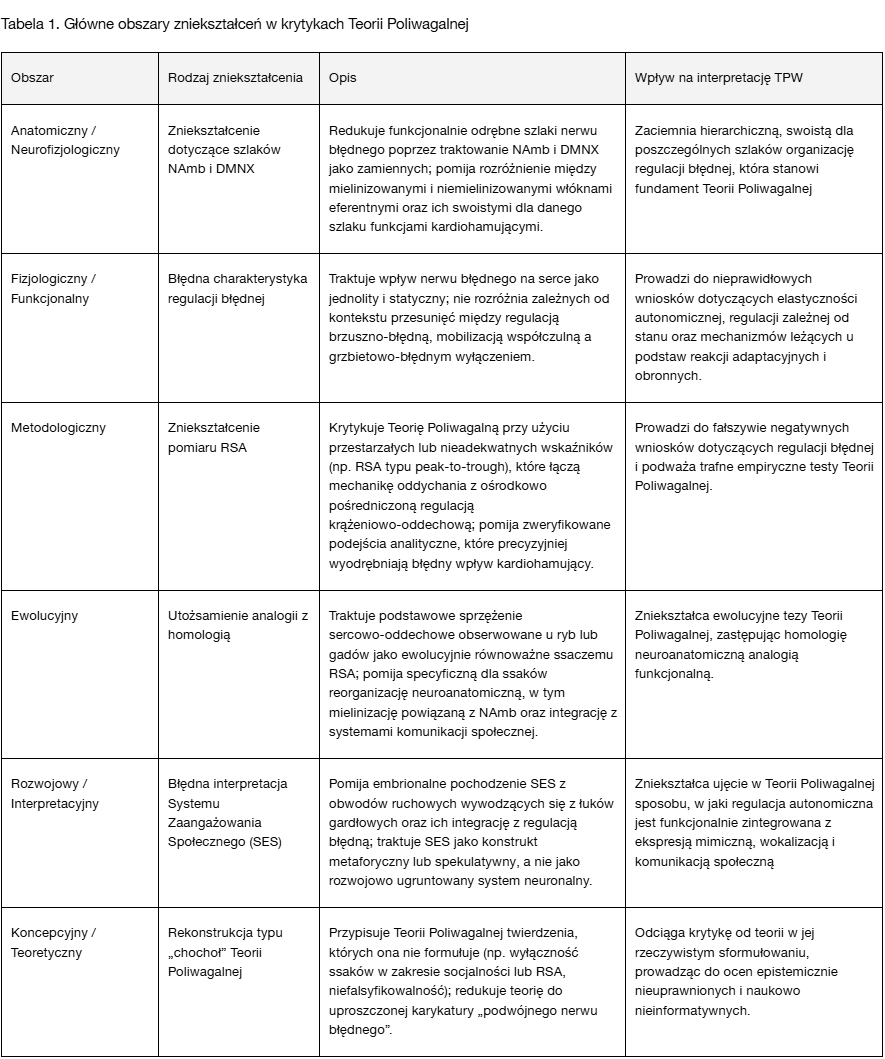
Jak pokazano w Tabeli 1, obszary te obejmują wymiary anatomiczne, fizjologiczne, ewolucyjne, koncepcyjne i metodologiczne. Choć treść poszczególnych krytyk bywa zróżnicowana, konsekwentnie mieszczą się one w tym typologicznym schemacie błędów — podważając spójność i naukową zasadność Teorii Poliwagalnej poprzez zniekształcanie jej podstawowych konstruktów lub stosowanie wadliwych kryteriów oceny. Ta uporządkowana rama dostarcza perspektywy diagnostycznej, umożliwiającej ocenę epistemicznej rzetelności zarówno krytyk akademickich, jak i kierowanych do szerokiego grona odbiorców.
Podczas gdy Tabela 1 wskazuje powtarzające się obszary błędów w różnych krytykach, Tabela 2 w bardziej szczegółowy sposób odnosi te zniekształcenia do kluczowych publikacji — pokazując, jak pierwotne błędne przedstawienia autorstwa Taylora ukształtowały wzorce, które następnie były powielane i rozwijane w późniejszych krytykach, zwłaszcza autorstwa Grossmana. Przywołania prac Taylora i jego współpracowników (np. Campbell i in., 2006; Monteiro i in., 2018; Sanches i in., 2019; Taylor i in., 2022) wskazują na powtarzalne schematy obecne w kolejnych publikacjach; konkretne dominujące typy błędów w poszczególnych artykułach przedstawiono w późniejszym zestawieniu typologicznym (zob. Tabela 4) oraz w bibliografii adnotowanej.
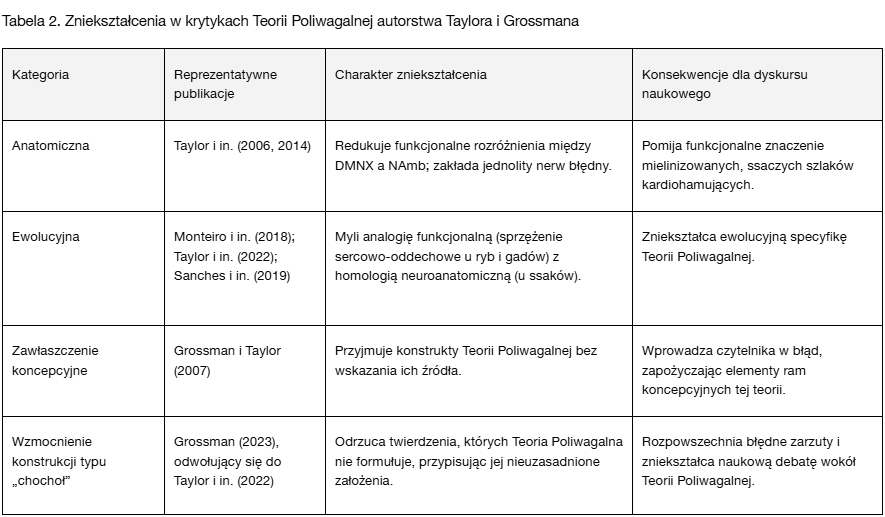
Jak pokazano w Tabeli 2, publikacje Taylora wielokrotnie redukują rozróżnienia anatomiczne oraz mylą konstrukty filogenetyczne stanowiące rdzeń Teorii Poliwagalnej. Późniejsze teksty Grossmana wzmacniają te zniekształcenia, osadzając je w krytykach o retorycznie autorytatywnym charakterze, lecz niezgodnych z danymi empirycznymi. To dziedziczenie błędów przyczynia się do wielowarstwowego zniekształcania Teorii Poliwagalnej zarówno w obszarze akademickim, jak i publicznym.
Podczas gdy Tabele 1 i 2 porządkują ogólne i fundamentalne zniekształcenia, Tabela 3 odnosi się bezpośrednio do głównych zarzutów formułowanych przez Grossmana — poddając je ocenie w świetle utrwalonych ustaleń empirycznych i neurofizjologicznych.
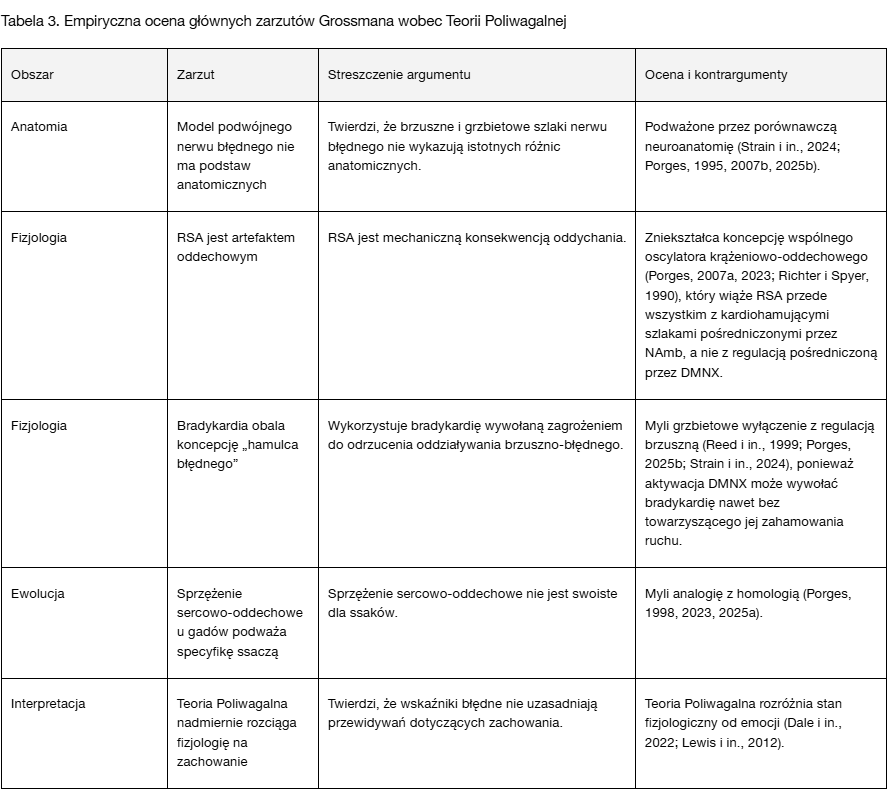
Tabela 3 zestawia główne twierdzenia wysuwane przez Grossmana (2007, 2023) i poddaje je ocenie w świetle współczesnych ustaleń empirycznych i neurofizjologicznych. W obszarach anatomicznym, fizjologicznym, ewolucyjnym i interpretacyjnym krytyki te konsekwentnie odzwierciedlają raczej nieporozumienie niż spór oparty na danych empirycznych — co wzmacnia potrzebę oceny zakorzenionej w literaturze pierwotnej.
Tabela 3 nie pełni wyłącznie funkcji podsumowania dowodów neuronaukowych podważających zarzuty Grossmana, lecz stanowi także ukazanie wewnętrznej spójności Teorii Poliwagalnej. Poprzez zgodność ze współczesnymi ustaleniami z zakresu neuroanatomii, fizjologii, psychobiologii rozwojowej i biologii ewolucyjnej teoria zachowuje integralność koncepcyjną i międzydyscyplinarną spójność — podkreślając swoją wyjaśniającą siłę, a nie podatność na redukcjonistyczną krytykę.
Rozpatrywane łącznie, omówione tu krytyki — czy to o charakterze anatomicznym, fizjologicznym, ewolucyjnym czy koncepcyjnym — mówią więcej o perspektywie interpretacyjnej ich autorów niż o zasadności samej Teorii Poliwagalnej. Utrzymujące się zniekształcenia, zwłaszcza w opracowaniach kierowanych do szerokiego grona odbiorców, podkreślają konieczność powrotu do źródeł pierwotnych przy ocenie twierdzeń tej teorii. Jak pokazuje niniejsza analiza, Teoria Poliwagalna pozostaje spójną i empirycznie zakorzenioną ramą teoretyczną, której międzydyscyplinarny zasięg nadal dostarcza wiedzy zarówno neuronauce podstawowej, jak i zastosowaniom translacyjnym.
Zniekształcenia metodologiczne i teza o artefaktowym charakterze RSA
Powtarzane przez Grossmana twierdzenie, że RSA jest jedynie artefaktem oddechowym, wynika z jego oparcia się na metodzie peak-to-trough w kwantyfikacji RSA — technice od dawna uznawanej za metodologicznie wadliwą. Po raz pierwszy skrytykowana przez Byrne’a i Porgesa (1993), a następnie potwierdzona przez Lewisa i in. (2012), metoda ta narusza podstawowe założenia statystyczne, jest nadmiernie wrażliwa na parametry oddechowe i trend sygnału oraz nie spełnia testów normalności i stacjonarności. W przeciwieństwie do niej metoda Porges–Bohrer wykazuje większą wrażliwość na blokadę błędną, jest mniej podatna na wpływ zmiennych oddechowych i lepiej nadaje się do prawidłowej analizy parametrycznej.
Pomimo wieloletniej krytyki Grossman nadal stosuje podejście peak-to-trough, co prowadzi do systematycznego obciążenia wyników i mylącej interpretacji związku RSA z brzusznym napięciem błędnym. Odmowa zmiany metod analitycznych lub uznania empirycznych obaleń sprawiła, że błąd ten utrzymuje się w publikacjach od ponad dwóch dekad.
W konsekwencji wiele opublikowanych krytyk Teorii Poliwagalnej nie opiera się na konkurencyjnych danych, lecz na pojedynczym, nieskorygowanym błędzie metodologicznym. Korekta tego rodzaju uchybień jest kluczowa dla przywrócenia rzetelności naukowej i odpowiedzialności analitycznej w dyskusji dotyczącej Teorii Poliwagalnej.
Czy Grossman krytykuje Teorię Poliwagalną, czy jej zniekształconą wersję?
Szczegółowa analiza krytyk Grossmana (2007, 2023) wskazuje, że jego argumenty często odnoszą się do wersji Teorii Poliwagalnej odbiegającej od jej sformułowania w literaturze pierwotnej. Rozbieżność ta wynika z selektywnego cytowania, odwoływania się do przestarzałych modeli fizjologii nerwu błędnego oraz błędnej interpretacji danych z zakresu neuroanatomii porównawczej. W rezultacie Teorii Poliwagalnej przypisywane są założenia, których ona nie formułuje (np. że RSA jest niezależne od oddychania lub że NAmb występuje wyłącznie u ssaków), a następnie stanowiska te są poddawane krytyce.
Ten schemat wskazuje na nieporozumienie o charakterze kategorialnym, a nie na spór oparty na konkurencyjnych danych empirycznych. W szczególności Grossman wielokrotnie utożsamia analogię funkcjonalną z homologią neuroanatomiczną, pomijając filogenetyczne uzasadnienie przedstawione w Teorii Poliwagalnej. Podobnie brak rozróżnienia między DMNX a NAmb zaciera anatomiczne podstawy hierarchicznego modelu regulacji autonomicznej proponowanego przez teorię.
Choć w tych krytykach często przywoływane są podstawowe publikacje Teorii Poliwagalnej, przedmiotem polemiki nie jest ich wierna reprezentacja ani oryginalne twierdzenia neurofizjologiczne teorii. Zamiast tego komentarze Grossmana opierają się na silnie zawężonym ujęciu wybranych subdyscyplin neuronauki — takich jak anatomia porównawcza czy neurofizjologia — bez odniesienia do integracyjnych modeli teoretycznych łączących strukturę, funkcję, rozwój i zachowanie. W konsekwencji krytyka koncentruje się nieproporcjonalnie na wtórnych interpretacjach i dalszych zastosowaniach klinicznych, zamiast na podstawowych konstruktach przedstawionych w literaturze źródłowej. Przedstawiana jako krytyka metodologiczna, ta linia argumentacji utrwala zniekształcenia pierwotnie wprowadzone przez Taylora i współpracowników, zastępując mechanizm metaforą oraz poddając ocenie wersję Teorii Poliwagalnej, która nigdy nie została sformułowana w jej źródłowych publikacjach.
Implikacje etyczne
Zniekształcenia Teorii Poliwagalnej wywołały szkody epistemiczne — wprowadzając w błąd klinicystów, dezorientując edukatorów oraz podważając przejrzystość translacyjną. Konsekwencje te nie mają wyłącznie charakteru akademickiego; utrudniają prawidłowe zastosowanie teorii opartej na zweryfikowanej neuroanatomii, rzetelnych wskaźnikach fizjologicznych oraz spójnym neuroetologicznym modelu regulacji autonomicznej.
Najnowsze ustalenia wzmacniają empiryczne podstawy Teorii Poliwagalnej. Na przykład Strain i in. (2024) wykazali, wykorzystując aktywację optogenetyczną, że neurony w DMNX — kluczowym elemencie grzbietowego kompleksu błędnego — wywołują bradykardię i modulują zachowania o charakterze lękowym bez wywoływania zahamowania ruchowego. Rozróżnienie to jest zgodne z rozróżnieniem w Teorii Poliwagalnej między reakcjami zagrożeniowymi pośredniczonymi przez system grzbietowy a immobilizacją bez strachu, odpowiadając na utrwalone nieporozumienia, w tym tych podtrzymywanych w numerze specjalnym Biological Psychology redagowanym przez Grossmana.
Kontekst ten rodzi dalsze wątpliwości natury etycznej. W numerze redagowanym gościnnie opublikowano wiele krytyk Teorii Poliwagalnej — takich jak artykuły Neuhubera i Berthouda (2022) oraz Doody’ego i in. (2023) — bez zapewnienia miejsca na odpowiedzi polemiczne lub reakcje naukowe. Biorąc pod uwagę ugruntowaną opozycję Grossmana wobec Teorii Poliwagalnej, brak stanowisk alternatywnych wprowadza możliwość stronniczości redakcyjnej. Taka asymetria niesie ryzyko zniekształcenia obrazu konsensusu naukowego oraz utrwalania błędów zarówno w środowisku akademickim, jak i w przestrzeni publicznej.
Jak zauważa Porges (2025b), krytyka naukowa powinna odnosić się do teorii w takiej postaci, w jakiej została ona sformułowana — a nie do jej karykaturalnych przedstawień. Etyczny dyskurs naukowy wymaga wierności dowodom, aktualności interpretacji oraz przejrzystości metodologicznej. Standardy te są niezbędne nie tylko dla rzetelnej oceny Teorii Poliwagalnej, lecz także dla zachowania integralności wymiany naukowej w szerszym sensie.
Zniekształcenia w dyskursie publicznym
Wiele zarzutów często przypisywanych Teorii Poliwagalnej — takich jak nadmierne uproszczenie czy nadmierna komercjalizacja — w istocie odnosi się do sposobu, w jaki teoria ta została przedstawiona lub zastosowana przez podmioty trzecie, a nie do jej wewnętrznych słabości.
- Nadmierne uproszczenie: Twierdzenia, że Teoria Poliwagalna upraszcza zależności między regulacją autonomiczną a emocjami, często wynikają z wtórnych interpretacji pomijających jej neurofizjologiczną złożoność. Konstrukty takie jak „hamulec wagalny (błędny)” czy „neurocepcja”, choć empirycznie uzasadnione, bywają w dyskursie popularnym redukowane do metafory. Uproszczenia te odzwierciedlają brak rzetelnego zapoznania się z literaturą pierwotną, a nie cechy immanentne Teorii Poliwagalnej.
- Nadmierna komercjalizacja: Teoria Poliwagalna bywa adaptowana w kontekstach coachingu, wellness czy działań brandingowych, które mogą nie spełniać standardów rygoru naukowego. Choć zjawisko to rodzi uzasadnione pytania dotyczące etyki translacyjnej, sama teoria zawiera w sobie rozważania etyczne i nie powinna być obarczana odpowiedzialnością za zniekształcenia wprowadzane przez podmioty zewnętrzne.
- Krytyka anatomicznego rozróżnienia między NAmb a DMNX: Neuhuber i Berthoud (2022) twierdzą, że proponowane przez Teorię Poliwagalną rozdzielenie anatomiczne nie znajduje potwierdzenia empirycznego. Ich krytyka pomija jednak dobrze ugruntowane dowody różnicujące NAmb i DMNX — obejmujące ich pochodzenie embriologiczne, mielinizację włókien eferentnych oraz swoistość funkcjonalną. Dane transkryptomiczne (Coverdell i in., 2019; Jalil i in., 2023) ujawniają markery molekularne swoiste dla neuronów NAmb, związane z funkcją autonomiczną i mielinizacją. Strain i in. (2024) wykazali ponadto, że selektywna aktywacja optogenetyczna neuronów DMNX wywołuje bradykardię i moduluje zachowania o charakterze lękowym, potwierdzając funkcjonalne i behawioralne znaczenie grzbietowego kompleksu błędnego w sposób opisany w Teorii Poliwagalnej.
- Krytyka filogenetyczna oparta na strukturach nerwu błędnego u gatunków innych niż ssaki: Niektóre zarzuty odwołują się do cech nerwu błędnego u ryb dwudysznych lub gadów, aby podważyć ewolucyjne twierdzenia Teorii Poliwagalnej. Argumenty te mylą analogię funkcjonalną z homologią filogenetyczną. Teoria Poliwagalna nie twierdzi, że regulacja błędna jest wyłączną cechą ssaków; wskazuje natomiast, że pośredniczony przez NAmb, mielinizowany komponent nerwu błędnego zintegrowany z czaszkowym systemem zaangażowania społecznego stanowi adaptację charakterystyczną dla ssaków. Utożsamianie ogólnej funkcji nerwu błędnego u różnych gatunków z tym wyspecjalizowanym systemem odzwierciedla niezrozumienie ewolucyjnej specyfiki Teorii Poliwagalnej.
Niezbędne jest rozróżnienie między empirycznymi podstawami teorii a jej publicznym przedstawieniem. Hasło w Wikipedii dotyczące Teorii Poliwagalnej stanowi przykład takiego pomieszania, często traktując niewłaściwe użycia lub zniekształcenia wprowadzane przez podmioty trzecie tak, jakby ujawniały one wady samej teorii. Stanowi to błąd kategorialny w ocenie naukowej.
Poddane uważnej analizie krytyki te nie ujawniają słabości teoretycznych, lecz trudności w rzetelnym upowszechnianiu wiedzy. Utrzymanie naukowej integralności Teorii Poliwagalnej wymaga precyzyjnej komunikacji oraz etycznej odpowiedzialności ze strony tych, którzy stosują ją w praktyce klinicznej, edukacyjnej i w przestrzeni publicznej
Typologia błędów w krytykach Teorii Poliwagalnej
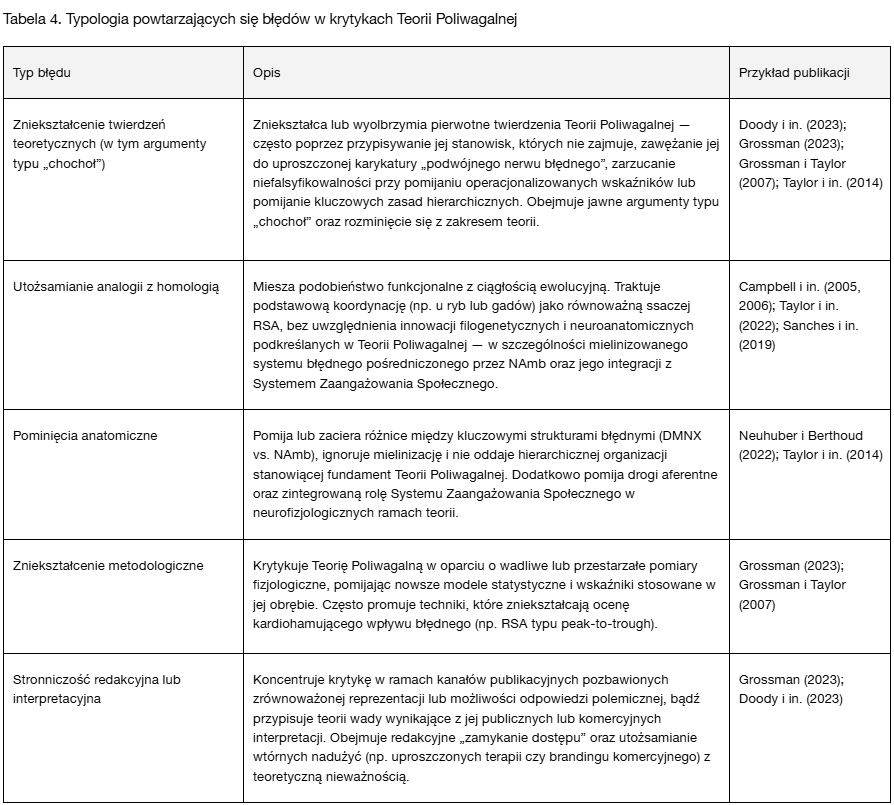
Aby syntetycznie ująć powtarzające się problemy zidentyfikowane w publikacjach krytycznych dotyczących Teorii Poliwagalnej, w Tabeli 4 przedstawiono typologię najczęściej występujących kategorii błędów. Obejmują one zarówno zniekształcenia retoryczne, jak i uchybienia empiryczne oraz metodologiczne. Każdy typ błędu został zdefiniowany, zilustrowany przykładami z opublikowanych krytyk oraz powiązany z wynikającym z niego zniekształceniem naukowym.
Ta klasyfikacja diagnostyczna podkreśla zasadniczą kwestię niniejszego opracowania: najbardziej utrwalone i widoczne krytyki Teorii Poliwagalnej często nie odnoszą się do niej w jej empirycznych i koncepcyjnych ramach. Zamiast przedstawiać ugruntowane wyzwania naukowe, stanowiska te nierzadko odzwierciedlają niezrozumienie biologii ewolucyjnej, neuroanatomii i poprawności metodologicznej — lub wynikają z uwarunkowań redakcyjnych i translacyjnych pozostających poza samą teorią.
Zakończenie
Kompleksowa analiza opublikowanych krytyk Teorii Poliwagalnej ujawnia powtarzalną strukturę błędu: najbardziej wpływowe zarzuty — w obszarach anatomicznym, fizjologicznym, ewolucyjnym i koncepcyjnym — wielokrotnie dają się powiązać z dorobkiem naukowym i wpływem redakcyjnym Paula Grossmana. Niezależnie od tego, czy poprzez bezpośrednie autorstwo, współpracę z E. W. Taylorem, czy redakcję numeru specjalnego Biological Psychology z 2023 roku, Grossman odegrał kluczową rolę w kształtowaniu dyskursu publicznego i akademickiego wokół Teorii Poliwagalnej.
Ta koncentracja sprzeciwu, zwłaszcza gdy jest wzmacniana przez formy publikacji wykluczające możliwość odpowiedzi polemicznych, rodzi uzasadnione wątpliwości dotyczące obiektywizmu naukowego oraz symulowania konsensusu. Krytyka naukowa powinna opierać się na wiernym przedstawieniu podstawowych twierdzeń teorii, a nie na ich karykaturalnym ujęciu czy retorycznym odwróceniu uwagi. Jak wykazano w niniejszym opracowaniu, wiele zarzutów wobec Teorii Poliwagalnej zniekształca jej kluczowe konstrukty, często poprzez niewielką liczbę błędów kategorialnych powtarzających się w różnych publikacjach i na różnych platformach.
Tego rodzaju powtarzalności nie należy utożsamiać ze zbieżnością stanowisk ani z konsensusem. Przeciwnie, wskazuje ona na potrzebę większej odpowiedzialności epistemicznej zarówno w środowisku akademickim, jak i w przestrzeni publicznej. Wzywa się czytelników i recenzentów do bezpośredniego sięgania do literatury pierwotnej, w której empiryczne podstawy i koncepcyjna architektura Teorii Poliwagalnej zostały przedstawione w najpełniejszy sposób.
Bibliografia:
Byrne, E. A., & Porges, S. W. (1993). Data-dependent peak–valley estimation of respiratory sinus arrhythmia: A methodological evaluation. Psychophysiology, 30(6), 672–678. https://doi.org/10.1111/j.1469-8986.1993.tb02061.x
Campbell, H. A., Taylor, E. W., & Egginton, S. (2005). Does respiratory sinus arrhythmia occur in fishes? Biology Letters, 1(4), 484–487. https://doi.org/10.1098/rsbl.2005.0365
Adnotacja: Utożsamianie analogii z homologią — Traktuje sprzężenie krążeniowo-oddechowe u ryb jako homologiczne wobec ssaczej RSA, myląc podobieństwo funkcjonalne z ciągłością filogenetyczną i pomijając oparte na NAmb, mielinizowane wyspecjalizowanie ssacze stanowiące centralny element Teorii Poliwagalnej.
Campbell, H. A., Leite, C. A. C., Wang, T., et al. (2006). Evidence for a respiratory component… in the rattlesnake. Journal of Experimental Biology, 209(14), 2628–2636. https://doi.org/10.1242/jeb.02278
Adnotacja: Utożsamianie analogii z homologią — Wykorzystuje występowanie sprzężenia podobnego do RSA u gadów, aby sugerować ewolucyjną równoważność z ssaczą RSA, pomijając nacisk Teorii Poliwagalnej na innowacje NAmb u ssaków oraz integrację z Systemem Zaangażowania Społecznego.
Coverdell, T., Ivison, R., Tao, J., and Campbell, J. (2019). 582-P: Disambiguating the nucleus ambiguus with single-cell transcriptomics. Diabetes 68 (Suppl. 1):582. https://doi.org/10.2337/db19-582-P
Dale, L. P., Kolacz, J., Mazmanyan, J., Leon, K. G., Johonnot, K., Bossemeyer Biernacki, N., & Porges, S. W. (2022). Childhood maltreatment influences autonomic regulation and mental health in college students. Frontiers in Psychiatry, 13, 841749. https://doi.org/10.3389/fpsyt.2022.841749
Doody, J. S., Burghardt, G. M., & Dinets, V. (2023). The evolution of sociality and the polyvagal theory. Biological Psychology, 180, Article 108569. https://doi.org/10.1016/j.biopsycho.2023.108569
Adnotacja: Zniekształcenie twierdzeń teoretycznych (w tym argumenty typu „chochoł”) — Przedstawia Teorię Poliwagalną jako twierdzącą, że gady są „aspołeczne”, oraz interpretuje jej argument o ssaczej innowacji autonomicznej jako zaprzeczenie socjalności gadów, myląc analogię behawioralną z neuroanatomicznym zakresem Teorii Poliwagalnej.
Grossman, P. (2023). Fundamental challenges and likely refutations of the five basic premises of the polyvagal theory. Biological Psychology, 180, 108589. https://doi.org/10.1016/j.biopsycho.2023.108589
Adnotacja: Zniekształcenie twierdzeń teoretycznych (argument typu „chochoł”) — Przypisuje Teorii Poliwagalnej twierdzenia, których ona nie formułuje (np. wyłączność ssaków w zakresie RSA). Zniekształca anatomiczną swoistość i fizjologiczne niuanse teorii. Nie uwzględnia odrębnych, kardiohamujących, mielinizowanych dróg jądra dwuznacznego (NAmb) oraz ich integracji z układami ruchowymi nerwów czaszkowych tworzącymi System Zaangażowania Społecznego, a także koordynującej roli wspólnego oscylatora krążeniowo-oddechowego w ssaczej regulacji autonomicznej.
Grossman, P., & Taylor, E. W. (2007). Toward understanding respiratory sinus arrhythmia: Relations to cardiac vagal tone, evolution and biobehavioral functions. Biological Psychology, 74(2), 263–285. https://doi.org/10.1016/j.biopsycho.2005.11.014
Adnotacja: Zniekształcenie twierdzeń teoretycznych (w tym argumenty typu „chochoł”) — Konstruuje argument typu „chochoł” wobec ewolucyjnych i dotyczących regulacji błędnej twierdzeń Teorii Poliwagalnej, przejmuje konstrukty Teorii Poliwagalnej bez właściwego przypisania autorstwa oraz zaciera rozróżnienie NAmb/DMNX, opierając się na nieprawidłowym ujęciu RSA.
Jalil, M., Coverdell, T. C., Gutierrez, V. A., Crook, M. E., Shi, J., Stornetta, D. S., Schwalbe, D. C., Abbott, S. B. G., & Campbell, J. N. (2023). Molecular disambiguation of heart rate control by the nucleus ambiguus [Preprint]. bioRxiv. https://doi.org/10.1101/2023.12.16.571991
Kolacz, J., Roath, O. K., Lewis, G. F., Karrento, K., et al. (2025). Cardiac vagal efficiency is enhanced by percutaneous auricular neurostimulation in adolescents with nausea: Moderation by antidepressant drug exposure. Neurogastroenterology & Motility, 37(5), e15007. https://doi.org/10.1111/nmo.15007
Lewis, G. F., Furman, S. A., McCool, M. F., & Porges, S. W. (2012). Statistical strategies to quantify respiratory sinus arrhythmia: A comparative evaluation. Biological Psychology, 89(2), 349–364. https://doi.org/10.1016/j.biopsycho.2011.11.009
Monteiro, D. A., Taylor, E. W., Sartori, M. R., Cruz, A. L., Rantin, F. T., & Leite, C. A. (2018). Cardiorespiratory interactions previously identified as mammalian are present in the primitive lungfish. Science Advances, 4(2), eaaq0800. https://doi.org/10.1126/sciadv.aaq0800
Adnotacja: Utożsamianie analogii z homologią — Wnioskuje o homologii ewolucyjnej na podstawie podobieństwa funkcjonalnego w interakcjach krążeniowo-oddechowych u ryb dwudysznych, pomijając podkreślane w Teorii Poliwagalnej swoiste dla ssaków przekształcenie neuroanatomiczne, w szczególności mielinizację powiązaną z NAmb oraz integrację z Systemem Zaangażowania Społecznego jako innowacje ewolucyjne.
Neuhuber, W. L., & Berthoud, H.-R. (2022). Functional anatomy of the vagus system: How does the polyvagal theory comply? Biological Psychology, 174, 108425. https://doi.org/10.1016/j.biopsycho.2022.108425
Adnotacja: Pominięcie anatomiczne — Redukuje Teorię Poliwagalną do uproszczonej narracji „podwójnego nerwu błędnego”, pomijając różnice rozwojowe, różnice w mielinizacji oraz hierarchiczne rozróżnienia między NAmb a DMNX. Ignoruje drogi aferentne oraz pomija ramy Systemu Zaangażowania Społecznego, przez co nie uwzględnia nacisku Teorii Poliwagalnej na zintegrowane obwody funkcjonalne i organizację ewolucyjną.
Sanches, P. V., Taylor, E. W., Durán, L. M., Cruz, A. L., Dias, D. P., & Leite, C. A. C. (2019). Respiratory sinus arrhythmia is a major component of heart rate variability in undisturbed, remotely monitored rattlesnakes (Crotalus durissus). Journal of Experimental Biology, 222, jeb197954. https://doi.org/10.1242/jeb.197954
Adnotacja: Utożsamianie analogii z homologią — Określa sprzężenie występujące u gadów jako RSA homologiczne wobec ssaczej RSA pośredniczonej przez NAmb, pomijając podkreślaną w Teorii Poliwagalnej swoistość ssaczej integracji neuroanatomicznej i behawioralnej.
Taylor, E. W., Campbell, H. A., Levings, J. J., Young, M. J., Butler, P. J., & Egginton, S. (2006). Coupling of the respiratory rhythm in fish with activity in hypobranchial nerves and with heartbeat. Physiological and Biochemical Zoology, 79(6), 1000–1009. https://doi.org/10.1086/507663
Adnotacja: Utożsamianie analogii z homologią — Interpretuje sprzężenie oddechowo-sercowe u ryb jako dowód ciągłości ewolucyjnej z ssaczą arytmią zatokową oddechową, nie odnosząc się do swoistych dla ssaków, mielinizowanych dróg błędnych powiązanych z NAmb oraz ich integracji z Systemem Zaangażowania Społecznego, podkreślanej w Teorii Poliwagalnej.
Taylor, E. W., Wang, T., & Leite, C. A. C. (2022). An overview of the phylogeny of cardiorespiratory control in vertebrates with some reflections on the “polyvagal theory.” Biological Psychology, 172, 108382. https://doi.org/10.1016/j.biopsycho.2022.108382
Adnotacja: Zniekształcenie twierdzeń teoretycznych (w tym argumenty typu „chochoł”) — Przedstawia Teorię Poliwagalną jako twierdzącą, że sprzężenie krążeniowo-oddechowe jest wyłączne dla ssaków, a następnie krytykuje to przeformułowane twierdzenie zamiast odnieść się do podkreślanego przez teorię znaczenia ssaczej specjalizacji neuroanatomicznej, w szczególności integracji brzusznego kompleksu błędnego wspierającej zaangażowanie społeczne.
Tutaj znajdziesz najnowsze artykuły naukowe dr Stephena Porgesa:
Wrzesień 2025: Polyvagal theory: a journey from physiological observation to neural innervation and clinical insight